The Challenge: Water for the right places at the right times
Our Australian waterbird populations depend on suitable feeding and nesting habitats coupled with flood events. Wetlands within the Murray-Darling Basin provide critical waterbird habitats, however the quality and availability of these sites are influenced by our management decisions. Protecting and maintaining suitable feeding and nesting habitats both between and during flood events is essential to maximise waterbird recruitment, maintain populations, and conserve biodiversity. This requires careful management of both vegetation and water regimes.
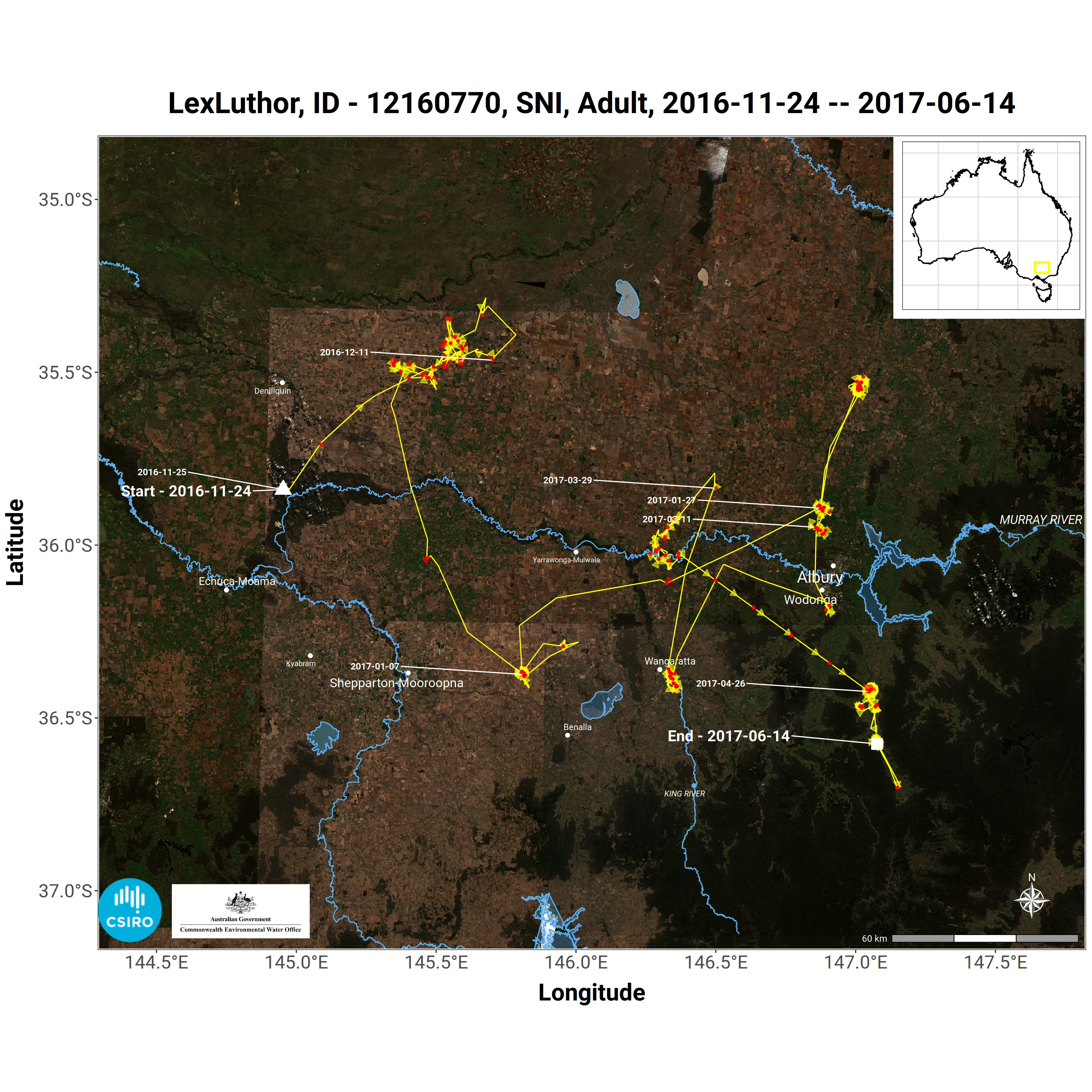
The timing and release of ‘environmental water’ within the Murray-Darling Basin is often focused on supporting successful completion of waterbird breeding events. Appropriately managing environmental water placement and timing is critical to facilitating the recruitment of juveniles into our waterbird populations and ultimately species survival. While some knowledge exists on the water flows needed to trigger waterbird nesting, we lack basic knowledge of how water flows interact with other factors such as predation and food abundance to influence waterbird breeding success and recruitment. We also lack knowledge of the foraging and dispersal movements of important species during and between breeding events – where do they go, and why? Filling these knowledge gaps is key to improving the efficiency of environmental water management – applying water to the right places at the right times – and ensuring the success of future breeding events and waterbird recruitment within the Basin.

A flock of straw-necked ibis (Threskiornis spinicollis) and Australian white ibis (Threskiornis molucca) take to the sky above their breeding site at Barmah-Millewa Forest, a wetland of the Murray River. Image credit: Heather McGinness
Our history
The Waterbird Research Project aims to fill knowledge gaps on waterbird ecology in the Basin to assist environmental water managers. From 2015-2019 the Waterbird Research Project was part of the Murray-Darling Basin Environmental Water Knowledge and Research (MDB EWKR) and was a collaboration between the CSIRO, University of NSW and University of Canberra. The MDB EWKR Project was funded by the Australian Government Commonwealth Environmental Water Office and managed by the Murray-Darling Freshwater Research Centre.
From 2019 we are continuing our research as part of the Flow-MER (Monitoring, Evaluation and Research) Program funded by the Commonwealth Environmental Water Office.
For more information on the Waterbird Research Project contact:
Dr Heather McGinness, Waterbird Research Project leader, CSIRO Land & Water
email: Heather.McGinness@csiro.au